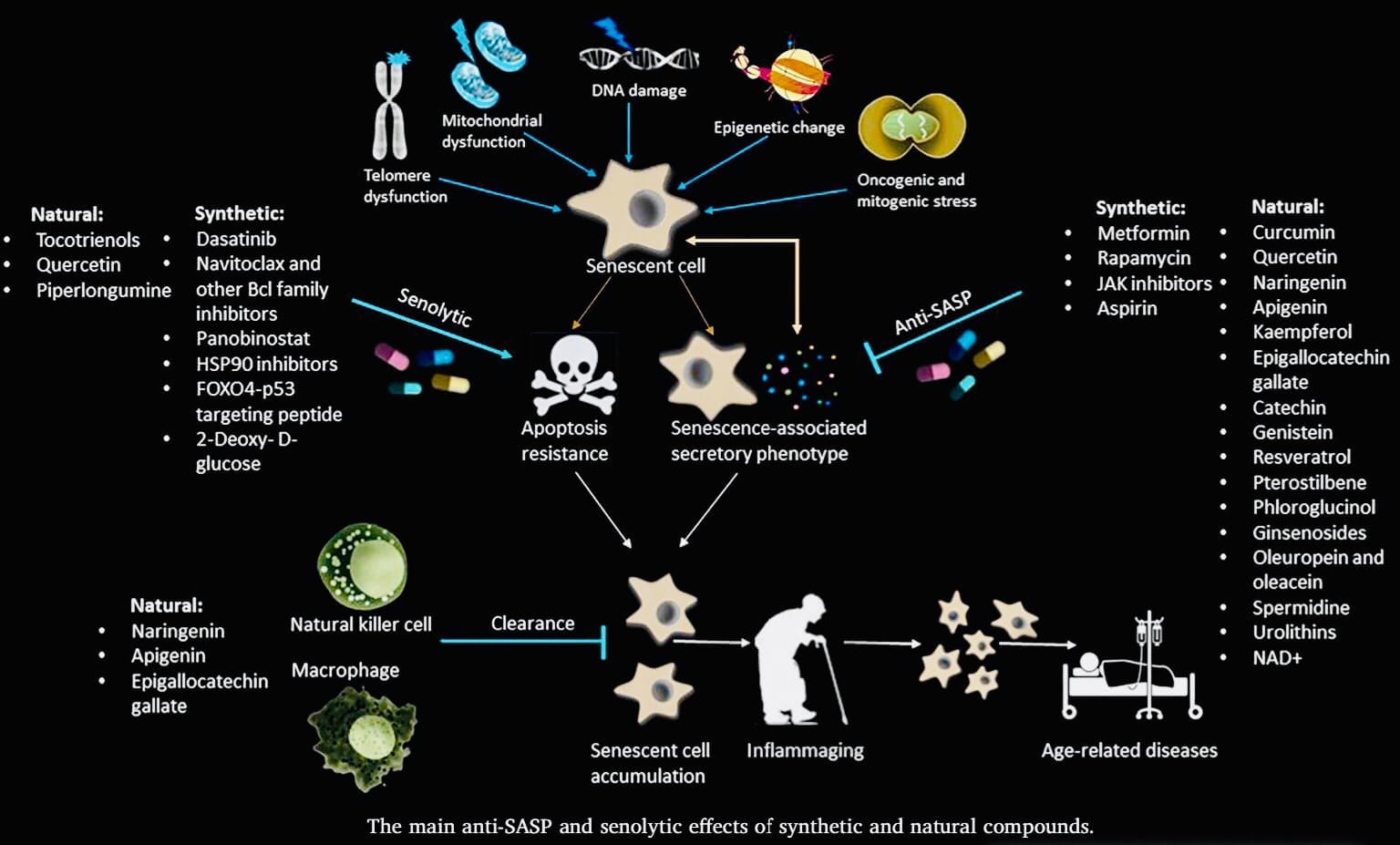
Senescent cells are like retired workers in our bodies. They’ve stopped dividing due to stress or damage, which prevents uncontrolled growth and potential cancer. However, these cells don’t disappear. They unleash a mix of signaling molecules called the Senescence-Associated Secretory Phenotype (SASP). While SASP can be helpful in wound healing,chronic SASP from accumulated senescent cells with age is linked to inflammation and age-related diseases.
This is where senolytics come in. These are drugs being developed to selectively target and eliminate senescent cells. By clearing these “troublemakers,” senolytics have the potential to improve healthspan and potentially lifespan by reducing chronic inflammation and delaying the onset of age-related diseases. Research in this area is ongoing, but the potential to combat aging and promote healthy longevity through senescent cell management is a promising avenue in human health.
Human adipocytes show a lower sensitivity to CD95-induced apoptosisthan preadipocytes.
The anti-apoptotic molecule Bcl-2 is up-regulated during adipogenic differentiation.
Overexpression of Bcl-2 rescues human preadipocytes from CD95-induced apoptosis.
Knockdown of Bcl-2 sensitizes human adipocytes to CD95-induced apoptosis.
Bcl-2 may constitute a new therapeutic target in the treatment of obesity.
Targeting apoptotic pathways in adipocytes has been suggested as a pharmacological approach to treat obesity. However, adipocyte apoptosis was identified as a cause for macrophage infiltration into adipose tissue. Previous studies suggest that mature adipocytes are less sensitive to apoptotic stimuli as compared to preadipocytes. Here, we aimed to identify proteins mediating apoptosis resistance in adipocytes.Our data revealed that the anti-apoptotic protein Bcl-2 (B-cell lymphoma 2) is up-regulated during adipogenic differentiation. Bcl-2 overexpression in preadipocytes lowers their apoptosis sensitivity to the level of mature adipocytes. Vice versa Bcl-2 knockdown in adipocytes sensitizes these cells to CD95-induced apoptosis. Taken together, our findings suggest a shift in the balance of pro-apoptotic and anti-apoptotic molecules during adipogenesis resulting in a higher apoptosis resistance. This study sheds new light on the apoptotic process in human fat cells and may constitute a new possible target for the specific regulation of adipose tissue mass.
Beyond a classical role in inhibiting apoptosis, BH4 domain has been characterized as a crucial regulator of other important cellular functions attributed to Bcl-2 and Bcl-xL, including proliferation, autophagy, differentiation, DNA repair, cell migration, tumor progression and angiogenesis.
The Bcl-2 family of proteins plays a central regulatory role in apoptosis. We have identified a novel, widely expressed Bcl-2 member which we have named Bcl-rambo. Bcl-rambo shows overall structural homology to the anti-apoptotic Bcl-2 members containing conserved Bcl-2 homology (BH) motifs 1, 2, 3, and 4. Unlike Bcl-2, however, the C-terminal membrane anchor region is preceded by a unique 250 amino acid insertion containing two tandem repeats. No interaction of Bcl-rambo with either anti-apoptotic (Bcl-2, Bcl-xL, Bcl-w, A1, MCL-1, E1B-19K, and BHRF1) or pro-apoptotic (Bax, Bak, Bik, Bid, Bim, and Bad) members of the Bcl-2 family was observed. In mammalian cells, Bcl-rambo was localized to mitochondria, and its overexpression induces apoptosis that is specifically blocked by the caspase inhibitors, IAPs, whereas inhibitors controlling upstream events of either the ‘death receptor’ (FLIP, FADD-DN) or the ‘mitochondrial’ pro-apoptotic pathway (Bcl-xL) had no effect. Surprisingly, the Bcl-rambo cell death activity was induced by its membrane-anchored C-terminal domain and not by the Bcl-2 homology region. Thus, Bcl-rambo constitutes a novel type of pro-apoptotic Bcl-2 member that triggers cell death independently of its BH motifs.
To remove senescent cells pharmacologically from non-genetically modified individuals, “senolytic” agents, including small molecules, peptides, and antibodies, are being developed21. Since the article describing the first senolytic agents was published in March, 201522, progress in identifying additional senolytic agents and their effects has been remarkably rapid. In that first article, a hypothesis-driven senolytic agent discovery paradigm was implemented. Senescent cells are resistant to apoptosis, despite the SASP factors they release, which should trigger apoptosis. Indeed, pro-apoptotic pathways are up-regulated in senescent cells22, yet these cells resist apoptosis23. The hypothesis was therefore tested that senescent cells depend on pro-survival pathways to defend against their own pro-apoptotic SASP. Using bioinformatics approaches based on the RNA and protein expression profiles of senescent cells, five Senescent-Cell Anti-Apoptotic Pathways (SCAPs) were identified (Table 1). That SCAPs are indeed required for senescent cell viability was verified by RNA interference studies, in which key proteins in these pathways were reduced. Through this approach, survival proteins were identified as the “Achilles’ heels” of senescent cells. Knocking-down expression of these proteins causes death of senescent but not non-senescent cells. Since the discovery of the first five SCAPs, another was identified (Table 1)24. This approach and the SCAPs discovered were subsequently used by others and us to identify putative senolytic targets22, 24, 25, 26, 27.
The SCAPs required for senescent cell resistance to apoptosis vary among cell types.The Achilles’ heels, for example of senescent human primary adipose progenitors differ from those in a senescent human endothelial cell strain, implying that agents targeting a single SCAP may not eliminate all types of senescent cells. So far, the senolytics that have been tested across a wide range of senescent cell types have all exhibited a degree of cell type specificity. For example, navitoclax is senolytic in a cell culture-acclimated human umbilical vein endothelial cell strain, but is not very effective against senescent primary human fat cell progenitors27. Even within a particular cell type, human lung fibroblasts, nativoclax is senolytic in the culture-acclimated IMR-90 lung fibroblast-like cell strain, while it is less so in primary human lung fibroblasts isolated from patients19, 27. Without extensive testing across a range of truly primary cells, as opposed to cell lines or culture-acclimated cell strains, it is difficult to contend that any particular candidate senolytic drug is universally effective for all types of senescent cells. Furthermore, senolytics can act synergistically in some cell types. For example, while neither dasatinib nor quercetin was significantly senolytic in mouse embryonic fibroblasts in vitro, the combination of dasatinib and quercetin was senolytic22. Thus, different senolytics may prove to be optimal for different indications and combinations of senolytics can be used to broaden the range of senescent cell types that are targeted.
YOU WANT TO INHIBIT THESE PATHWAYS TO KILL SENESCENT CELLS:
Increases in cell size, lipofuscin accumulation, high expression of the cell cycle regulator, p16INK4A, p21CIP1, and SASP factors (e.g., IL-6, IL-8, monocyte chemoattractant protein-1, plasminogen-activated inhibitor-1, and many others), increased cellular senescence-associated β-galactosidase (SA-βgal) activity, and appearance of senescent-associated distension of satellites (SADS) and telomere-associated DNA damage foci (TAFs), among others. None of these markers are fully sensitive or specific, so combinations of them are needed to draw conclusions about effects of diseases or interventions on senescent cell numbers.
Cellular senescence is a tumor-suppressive mechanism that permanently arrests cells at risk for malignant transformation. However, accumulating evidence shows that senescent cells can have deleterious effects on the tissue microenvironment. The most significant of these effects is the acquisition of a senescence-associated secretory phenotype (SASP) that turns senescent fibroblasts into proinflammatory cells that have the ability to promote tumor progression.
The tissue microenvironment is defined by the phenotypes of the cells in the immediate area and by the physical and chemical properties of the soluble and insoluble factors surrounding cells within a given tissue. These properties include temperature and oxygen tension, as well as various molecules that may be produced locally—for example, growth factors and cytokines. Further, cells within tissues form a dynamic network that contributes to their microenvironment. At the same time, the tissue microenvironment regulates cell behavior. This reciprocal relationship determines tissue function and repair and is also central to a number of pathologies, including cancer.
A permissive microenvironment supports and promotes tumor growth and cancer cell aggressiveness (1–4). Alterations in the cellular and molecular composition of the connective tissues surrounding carcinomas allow tumors to evade detection by the immune system as well as to proliferate inappropriately, invade the surrounding tissue structure, and eventually metastasize. The synergy between an altered microenvironment and the genetic alterations acquired by tumor cells allows these cells to evade preventive mechanisms and become fully malignant. Cellular senescence is now recognized as a potent tumor-suppressive mechanism that arrests the growth of cells at risk for malignant transformation (5–12). However, recent studies show that senescent cells develop altered secretory activities that may induce changes in the tissue microenvironment, relaxing its control over cell behavior and promoting tumorigenesis (13–18).
How can the senescence response be both tumor suppressive and procarcinogenic? It is important to consider that a biological process such as cellular senescence can be both beneficial and deleterious. The idea that processes can have such dual effects is consistent with a major evolutionary theory of aging termed antagonistic pleiotropy (19). The senescence-associated secretory phenotype (SASP) represents one of the darkest sides of the senescence response and is the focus of this review. We particularly emphasize the potential effects of the SASP (20) on cell behavior in the context of tumor progression.
CELLULAR SENESCENCE
Cellular senescence occurs in culture and in vivo as a response to excessive extracellular or intracellular stress. The senescence program locks the cells into a cell-cycle arrest that prevents the spread of damage to the next cell generation and precludes potential malignant transformation (19). Senescent cells have been shown to accumulate over the life span of rodents, nonhuman primates, and humans (21). These cells are found primarily in renewable tissues and in tissues that experience prolonged inflammation.
A plethora of stresses can provoke cellular senescence (22, 23). These stresses include telomeric dysfunction (telomere uncapping) resulting from repeated cell division (termed replicative senescence), mitochondrial deterioration, oxidative stress, severe or irreparable DNA damage and chromatin disruption (genotoxic stress), and the expression of certain oncogenes (oncogene-induced senescence) (see Figure 1) (24–31). Stresses that cause cellular senescence can be induced by external or internal chemical and physical insults encountered during the course of the life span, during therapeutic interventions (for example, X-irradiation or chemotherapy), or as a consequence of endogenous processes such as oxidative respiration and mitogenic signals. External mitogenic signals, for example growth-related oncogene alpha (GROα) secretion by tumor cells in close proximity to normal cells (32) or circulating angiotensin II (33, 34), have also been shown to induce cellular senescence. All somatic cells that have the ability to divide can undergo senescence. Regardless of the disparate mechanisms of senescence-inducing stresses, the senescence program is activated once a cell has sensed a critical level of damage or dysfunction. So far, the senescence growth arrest has been shown to depend on the activities of the major tumor-suppressor pathways controlled by p16INK4a and pRB (retinoblastoma protein), as well as by p53. Some of the molecules involved in pathways upstream and downstream of the senescence-associated phenotype have been used as markers to detect senescent cells in culture and in vivo.
Multiple types of stimuli can provoke cellular senescence and a senescence-associated secretory phenotype (SASP). When irreversible cell-cycle arrest is triggered by severe DNA damage (i.e., dysfunctional telomeres or oncogenic stress), the SASP occurs…
THE SECRETORY PHENOTYPE OF SENESCENT CELLS
The senescent phenotype is not limited to an arrest of cell proliferation. In fact, a senescent cell is a potentially persisting cell that is metabolically active and has undergone widespread changes in protein expression and secretion, ultimately developing the SASP. This phenotype has also been termed the senescence-messaging secretome (35). We recently provided a large-scale characterization of the SASP, using antibody arrays to quantitatively measure factors secreted by human fibroblasts and epithelial cells (18), as well as mouse fibroblasts (J.P. Coppé & J. Campisi, unpublished data). The potential existence of the SASP was already suggested by large-scale comparative gene (mRNA) expression studies performed on fibroblasts from different-aged donors and different tissues of origin (36–46). Among the cells that have been shown to senesce and secrete biologically active molecules are liver stellate cells (47), endothelial cells (36, 48–51), and epithelial cells of the retinal pigment, mammary gland, colon, lung, pancreas, and prostate (8, 18, 36, 41, 52–56).
Senescence-associated changes in gene expression are specific and mostly conserved within individual cell types. Most differences between the molecular signatures of presenescent and senescent cells entail cell-cycle- and metabolism-related genes, as well as genes encoding the secretory proteins that constitute the SASP. The SASPincludes several families of soluble and insoluble factors (see Table 1). These factors can affect surrounding cells by activating various cell-surface receptors and corresponding signal transduction pathways that may lead to multiple pathologies, including cancer. SASPfactors can be globally divided into the following major categories: soluble signaling factors (interleukins, chemokines, and growth factors), secreted proteases, and secreted insoluble proteins/extracellular matrix (ECM) components. SASP proteases can have three major effects: (a) shedding of membrane-associated proteins, resulting in soluble versions of membrane-bound receptors, (b) cleavage/degradation of signaling molecules, and/or (c) degradation or processing of the ECM. These activities provide potent mechanisms by which senescent cells can modify the tissue microenvironment. In the following sections, we discuss these SASP subsets and some of their known paracrine effects on nearby cells, with an emphasis on their ability to facilitate cancer progression.
Table 1 The senescence-associated secretory phenotype (SASP). Factors significantly altered between presenescent and senescent states are listed.
Soluble Signaling Factors as Major Components of the Senescence-Associated Secretory Phenotype
Senescent cells secrete interleukins, inflammatory cytokines, and growth factors that can affect surrounding cells.
IL-6
The most prominent cytokine of the SASP is interleukin-6 (IL-6), a pleiotropic proinflammatory cytokine(see Figure 2). IL-6 has been shown to be associated with DNA damage– and oncogenic stress–induced senescence of mouse and human keratinocytes, melanocytes, monocytes, fibroblasts, and epithelial cells (16, 18, 57, 58). Further, IL-6 secretion appears to be directly controlled by persistent DNA-damage signaling (ATM and CHK2), independent of the p53 pathway (59). Through IL-6 expression, senescent cells can directly affect neighboring cells that express the IL-6R (gp80) and gp130 signaling complex at their surface, such as epithelial and endothelial cells of various functions and origins.
Human fibroblasts, either presenescent (PRE) or senescent (SEN), were immunostained for the inflammatory cytokines interleukin (IL)-6 and IL-8, as well as the senescence marker p16. Cells were made senescent either by replicative exhaustion (REP) or ionizing …
IL-1
Another interleukin signaling pathway demonstrated to be upregulated by senescent cells is that of IL-1(60, 61). Both IL-1α and -β are overexpressed and secreted by senescent endothelial cells (62), fibroblasts (63, 64), and chemotherapy-induced senescent epithelial cells (53). These cytokines can affect neighboring cells through the cell-surface receptors (IL-1 receptor/Toll-like receptor superfamily), which act primarily to trigger the nuclear factor kappa B and activating protein 1 pathways (65).
Chemokines (CXCL and CCL)
Most senescent cells overexpress IL-8 (CXCL-8) (see Figure 2), along with GROα and GROβ (CXCL-1 and -2; the murine CXCL-1 is named KC) (58, 66, 67). CCL family members that are generally upregulated in senescent cells include MCP-2, -4, and -1 (CCL-8, -13, and -2); HCC-4 (CCL-16); eotaxin-3 (CCL-26); and macrophage inflammatory protein (MIP)-3α and -1α (CCL-20, -3). MCP-3 (CCL-7) is overexpressed by senescent liver stellate cells and by prostate and skin fibroblasts. Fibroblasts induced to senesce by oncogenic RAS secrete high levels of MCP-3 as well as I-309 (CCL-1). In addition, both fibroblasts induced to senesce by RAS and stellate cells induced to senesce by liver damage secrete high levels of another two members of the CXCL family, GCP-2 (CXCL-6) and ENA-78 (CXCL-5). Overexpression of PF-4 (CXCL-4) and SDF-1 (CXCL-12) was observed in senescent prostate fibroblasts (46, 68). Recently, it was shown that cells undergoing oncogene-induced senescence secrete multiple CXCR-2 (IL-8RB)-binding chemokines (15). It was proposed that senescent cells activate a self-amplifying secretory network in which CXCR-2-binding chemokines reinforce growth arrest.
IGF pathway
The insulin-like growth factor (IGF)/IGF receptor network may also contribute to the effect senescent cells exert on their microenvironment. Senescent endothelial, epithelial, and fibroblast cells express high levels of almost all the IGF-binding proteins(IGFBPs), including IGFBP-2, -3, -4, -5, and -6 (18, 69, 70) and their regulators, IGFBP-rP1 and -rP2 [also known as connective tissue growth factor (CTGF)] (44, 71). Recently, activation of the BRAF oncogene in primary fibroblasts was shown to lead to the secretion of IGFBP-7, which acts through autocrine/paracrine pathways to induce senescence and apoptosis in neighboring cells (72).
Other soluble factors
There are additional soluble factors associated with the SASP. For example, inflammatory cytokines such as the colony-stimulating factors (CSFs, including GM-CSF and G-CSF) are secreted at high levels by senescent fibroblasts (18). In addition, osteoprotegerin, a secreted decoy receptor for tumor necrosis factor alpha, is present at high levels in the extracellular milieu of senescent fibroblasts. Other molecules upregulated at senescence include prostaglandin E2 (PGE2) (57, 73) and Cox-2,the enzyme responsible for the production of PGE2 and other prostaglandins.
EFFECTS ON CELL BEHAVIOR
Factors secreted by senescent cells can promote tumor development in vivo and malignant phenotypes such as proliferation and invasiveness in cell-culture models. These effects have been observed in a number of tissues, including breast (13, 18, 77, 78, 114), skin (115), prostate (18, 116), pancreas (117), and oropharyngial mucosa (14). The effects of the complex SASP are, of course, dependent on the tissue context. Thus, different models show different effects of the SASP. In the following sections, we discuss in greater detail the various behavioral changes cells can undergo when residing in the proximity of senescent cells and how the senescent tissue microenvironment can facilitate tumor initiation and progression.
Cellular senescence was first described as a failure of normal human cells to divide indefinitely in culture. Until recently, the emphasis in the study of cell senescence has been focused on the accompanying intracellular processes. The focus of the attention has been on the irreversible growth arrest and two important physiological functions that rely on it: suppression of carcinogenesis due to the proliferation loss of damaged cells, and the acceleration of organism aging due to the deterioration of the tissue repair mechanism with age.
However, the advances of the past years have revealed that senescent cells can impact the surrounding tissue microenvironment, and, thus, that the main consequences of senescence are not solely mediated by intracellular alterations. Recent studies have provided evidence that a pool of molecules secreted by senescent cells, including cytokines, chemokines, proteases and growth factors, termed the senescence-associated secretory phenotype (SASP), via autocrine/paracrine pathways can affect neighboring cells. Today it is clear that SASPfunctionally links cell senescence to various biological processes, such as tissue regeneration and remodeling, embryonic development, inflammation, and tumorigenesis. The present article aims to describe the “social” life of senescent cells: basically, SASPconstitution, molecular mechanisms of its regulation, and its functional role.
It is well known that the main features of CS are similar across its different forms and different types of proliferating cells [40]. Figure 2 shows the most important “individual” intracellular changes that accompany CS, which are subdivided into events occurring in the nucleus and in the cytoplasm. The change in the secretory profile occupies a special place among the modifications accompanying CS. It is generally accepted that the senescence– associated secretory phenotype (SASP) defines the engagement of senescent cells in a wide range of processes, such as reparation, propagation of senescence, immune clearance, embryogenesis, and tumorigenesis [29, 31, 38, 79, 80].
Classification of SASP factors
The term SASP was first used in 2008 to refer to the factors secreted by senescent cells [24]. The following classification of SASP components has been adopted: soluble signaling factors, proteases, insoluble extracellular matrix proteins, and non-protein components [78]. SASP factors can be divided into the following groups based on molecular mechanisms [81]:
1) Factors binding to a receptor. This group includes soluble signaling molecules, such as cytokines, chemokines, and growth factors. These factors can influence cells of the microenvironment by interacting with the corresponding surface receptors on their membranes and, thus, triggering various intracellular signaling cascades [82, 83]. The most well known representatives of this group are interleukins IL-6, IL-8, IL-1a, chemokines GROα, GROβ, CCL-2, CCL-5, CCL-16, CCL-26, CCL-20, and the growth factors HGF, FGF, TGFβ, and GM-CSF.
2) Factors acting directly. This group includes matrix metalloproteases MMP-1, MMP-10, MMP-3 and serine proteases: the tissue plasminogen activator (tPA) and urokinase plasminogen activator (uPA). These factors are capable of cleaving membrane-bound proteins, destroying signaling molecules and remodeling the extracellular matrix, to enable senescent cells to modify their microenvironment [84]. Small non-protein components, such as reactive oxygen (ROS) and nitrogen species that damage neighboring cells, can also be included in this group [78, 85].
3) Regulatory factors. This group includes tissue inhibitors of metalloproteases (TIMP), the plasminogen activator inhibitor (PAI), and insulin-like growth factor binding proteins (IGFBP). These factors do not have their own enzymatic activity. However, when they bind to factors from the first and second groups, they regulate their functioning. For example, TIMP inhibits the activity of most MMPs [86], PAI-1 functions primarily as an inhibitor of tPA and uPA [87], and IGFBP function as IGF transport proteins [88].
In addition to all the factors mentioned above, which are secreted by senescent cells, another component has recently begun to be viewed as part of SASP: extracellular vesicles, in particular vesicles associated with microRNAs [89]. It turns out that such vesicles can affect neighboring cells and cells located at a considerable distance, both by initiating and suppressing CS, depending on the composition of microRNAs.
It should be emphasized that the specific qualitative and quantitative composition of the secreted factors largely depends on the type of cells and the inducer of senescence, which makes it very difficult to study this CS feature. Several approaches to the study of SASP and elucidation of the functions of its individual components have been described to date. The main approaches are presented at Fig. 3.
Experimental approaches to study SASP and to identify the functional role of its individual components
Mechanisms of SASP regulation
It is well known that cellular senescence is not a onetime phenomenon, but one that develops over time [99]. Remarkably, SASP has also recently begun to be viewed as a dynamic process which can be subdivided into several phases [16]. It is believed that the first phase of secretion begins immediately after DNA damage and lasts for the first 36 hours. It should be noted that the onset of this phase is not sufficient evidence in favor of initiation of senescence, since it does not preclude complete repair or apoptosis [99]. The next phase is the “early” SASP phase, which continues for several days after the initiation of CS. It is during this period that the most important SASP factors, for example IL-1α, start to appear. During the next 4–10 days, the secretion of most factors intensifies due to the autocrine effect of SASP, which ultimately leads to the formation of “mature” SASP [16]. Such a wave-like secretion of factors during the development of CS is largely attributed to positive feedback loops and complex regulatory mechanisms. The most common mechanisms for SASP regulation are presented below.
It should be noted that SASP is regulated both at the transcriptional and post-transcriptional levels. The key role in the regulation of SASP components expression, including IL-6, IL-8, CXCL1, and CXCR2, belongs to the nuclear factor kappa-light-chain-enhancer of activated B cells, NF-kB [100–102]. For most of these factors, control over transcription is achieved through positive feedback loops. A vivid example of such “self-amplifying” loops is the regulation of IL-1α secretion [15, 103]. It has been reported that another transcription factor, C/EBPβ, is also involved: by binding directly to the promoter of the IL-6 gene, where it initiates its expression [82, 104].
At the post-transcriptional level of SASP regulation, it is customary to identify DDR (DNA Damage Response)-dependent and independent mechanisms [15]. As mentioned above, one of the most important features of CS is the DNA damage response. It has been shown that knockdowns of such DDR components as ATM, Chk2, NBS1, and H2AX reduce the expression and, accordingly, the secretion of a number of SASP factors, including IL-6 and IL-8 [104–106]. Despite evidence that DDR is involved in SASP regulation, the detailed mechanisms for their relationships are not fully understood. The signaling pathways known today are associated with the ability of DDR components, in particular ATM kinase, to somehow regulate NFkB activity. For example, ATM can form complexes with the NEMO protein, which, due to the initiation of DDR, are exported from the nucleus to the cytoplasm, where NEMO binds to and activates IKK kinase. IKK promotes the dissociation of the inhibitory IkB protein from its complex with NF-kB and activation of the latter [107].
More recently, the involvement of the transcription factor GATA4 in the DDR-dependent mechanism of SASPregulation has been demonstrated [108]. Normally, GATA4 is degraded by p62-mediated autophagy. However, autophagy is suppressed in most senescent cells, and, therefore, GATA4 stabilizes, and this process is ATM-dependent. The accumulation of GATA4 in senescent cells facilitates the initiation and maintenance of NF-kB activity.
List of markers/methods for senescent cell detection: Marker/Method
Large and flat morphology (Various microscopical and staining approaches)
Lack of cell proliferation markers: absence of Ki-67, BrdU/EdU-incorporation, no colony formation
Lack of response to growth signals
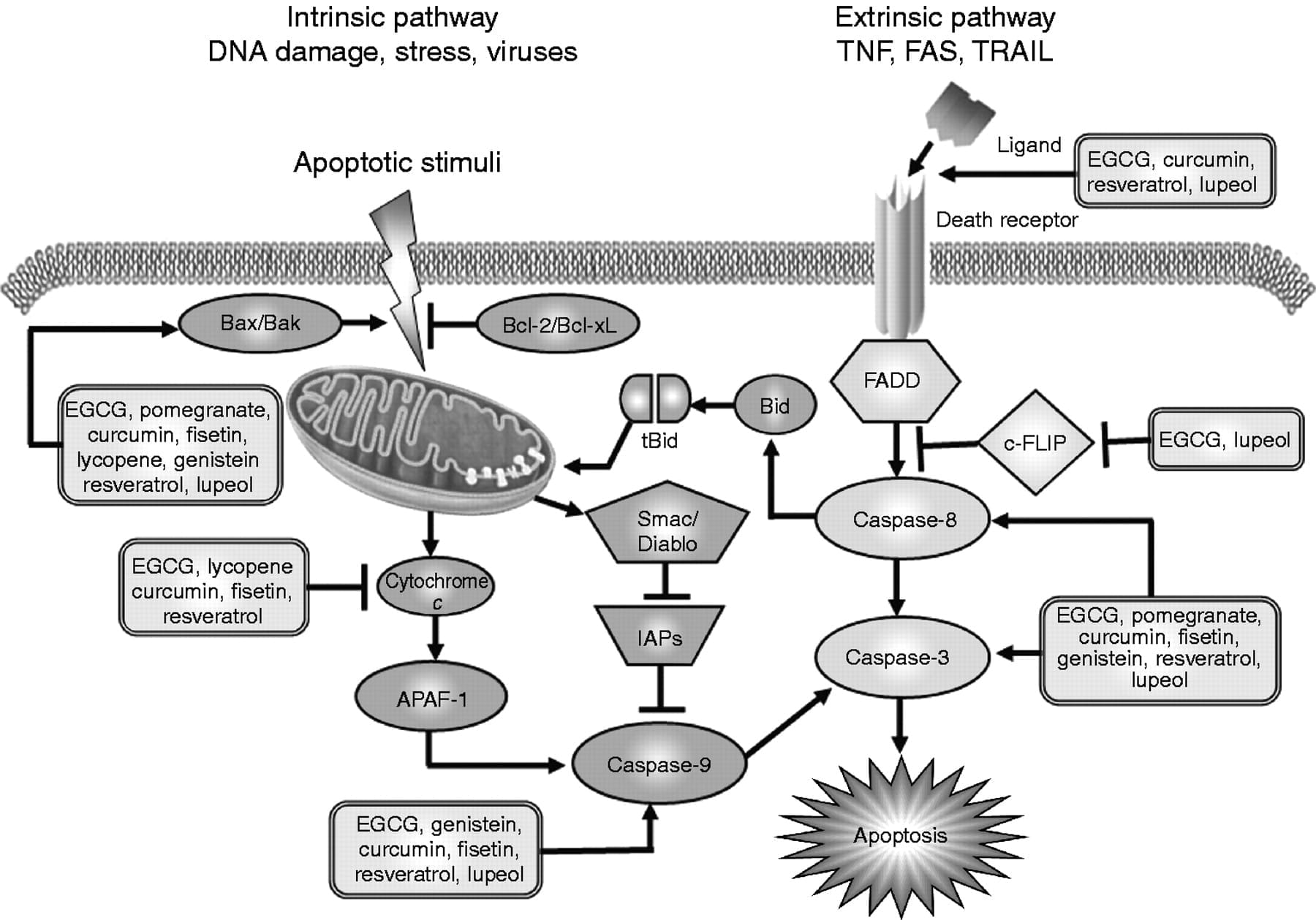
Resistance to apoptosis: BCL family members ( Bcl-2, Bcl-w, or Bcl-xL)
H Thoppil, K Riabowol – Frontiers in Cell and Developmental Biology, 2019
… Huang et al., 1999; Montero et al., 2011). These compounds form one of the first discovered members of the senolytic class of drugs that selectively induce apoptosis in senescent cells. Four years after their initial identification as …
M Demaria – US Patent App. 16/492,410, 2020
… 15. The drug conjugate according to claim 12, wherein the cytotoxic agent is a senolytic agent, a radioisotope, a toxin or a toxic peptide. 16 … The cytotoxic agent can be a radioisotope, a toxin, toxic peptide or a senolytic drug …
ZN Wang, RN Su, BY Yang, KX Yang, LF Yang, Y Yan… – Frontiers in Cell and …, 2020
… proteases expressed by senescent lung fibroblasts could result in low-level inflammation and fibrosis (Schafer et al., 2017; Álvarez et al., 2017). Clearance of these senescent fibroblasts by senolytic drugs would render the reso.
LITO Salon
… However, there is no single senolytic that has been shown to target all of our senescent cells … One example is Cleara Biotechnologies, whose founder, Dr. Peter De Keizer, has talked about senolytic “cocktails” and the problem …
R Sharma, Y Padwad – Trends in Food Science & Technology, 2020
JavaScript is disabled on your browser. Please enable JavaScript to use all the features on this page. Skip to main content Skip to article …
BA Benayoun, RA Veitia
… aging. From the perspective of precision medicine, it will be possible to identify individu- als at higher risk of developing single or multiple morbidi- ties for devising new preventive measures, such as specific senolytic treatments …
S Saxena, S Kumar – GeroScience
… 2017). Senescent cells also upregulate the expression of anti-apoptotic proteins BCL-W and BCL-XL, the inhibition of which with siRNA or small molecules ABT-737 or ABT-263 (senolytic drugs) leads to apoptosis (Chang …
A Costa, MY Bonner, S Rao, L Gilbert, M Sasaki… – Cancers, 2020
… the nonpermissive temperature. This suggests that the maintenance of the senescent phenotype results in a low level of p38 signaling and that p38 activators might be effective in mediating senolytic therapies. Consistent with …
E Laconi, F Marongiu, J DeGregori – British Journal of Cancer, 2020
Why do we get cancer mostly when we are old? According to current paradigms, the answer is simple: mutations accumulate in our tissues throughout life, and some of these mutations contribute to cancers.
R Kumar, A Sharma, Y Padwad, R Sharma – Biogerontology
… Working on green tea EGCG, we have previously reported its anti- immunosenescence, anti-inflammatory, anti-senescence, senolytic and synbiotic attributes thereby suggesting its multi-faceted potency as an …
Q Li – 2020
Page 1. From the Department of Cell and Molecular Biology Karolinska Institutet, Stockholm, Sweden Novel properties of mature adipocytes in obesity and hyperinsulinemia Qian Li 李 倩 Stockholm 2020 Page 2. Cover picture …
A Costa, MY Bonner, S Rao, L Gilbert, M Sasaki… – Cancers, 2020
… the nonpermissive temperature. This suggests that the maintenance of the senescent phenotype results in a low level of p38 signaling and that p38 activators might be effective in mediating senolytic therapies. Consistent with …
E Laconi, F Marongiu, J DeGregori – British Journal of Cancer, 2020
Why do we get cancer mostly when we are old? According to current paradigms, the answer is simple: mutations accumulate in our tissues throughout life, and some of these mutations contribute to cancers. Although …
R Kumar, A Sharma, Y Padwad, R Sharma – Biogerontology
… Working on green tea EGCG, we have previously reported its anti- immunosenescence, anti-inflammatory, anti-senescence, senolytic and synbiotic attributes thereby suggesting its multi-faceted potency as an …
Q Li – 2020
Page 1. From the Department of Cell and Molecular Biology Karolinska Institutet, Stockholm, Sweden Novel properties of mature adipocytes in obesity and hyperinsulinemia Qian Li 李 倩 Stockholm 2020 Page 2. Cover picture …
A Chandra, AB Lagnado, JN Farr, DG Monroe, S Park… – Journal of bone and mineral …, 2020
… To test if senolytic drugs, which clear senescent cells, alleviate FRT-related bone damage, we administered the senolytic agents, Dasatinib (D), Quercetin (Q), Fisetin (F), and a cocktail of D and Q (D+Q). We found moderate …
RM Laberge, J Campisi, M Demaria, N David… – US Patent App. 16/584,638, 2020
… US 20200030323 A1 US20200030323 A1 US 20200030323A1 US 201916584638 A US201916584638 A US 201916584638A US 2020030323 A1 US2020030323 A1 US 2020030323A1 Authority US United States Prior …
T Montero-Melendez, A Nagano, C Chelala, A Filer… – Nature Communications, 2020
Rheumatoid arthritis affects individuals commonly during the most productive years of adulthood. Poor response rates and high costs associated with treatment mandate the search for new therapies. Here we show that targeting a specific …
CL Xu, B Sang, GZ Liu, JM Li, XD Zhang, LX Liu… – Nucleic Acids Research, 2020
… Moreover, SENEBLOC was shown to be involved in both oncogenic and replicative senes- cence, and from the perspective of senolytic agents we show that the antagonistic actions of rapamycin on senescence are dependent on SENEBLOC ex- pression …
SS Çinarouglu, E Timucin, GB Akcapinar, U Sezerman… – US Patent App. 16/526,997, 2020
… US2020031873A1 US 20200031873 A1 US20200031873 A1 US 20200031873A1 US 201916526997 A US201916526997 A US 201916526997A US 2020031873 A1 US2020031873 A1 US 2020031873A1 …
A Barbouti, PVS Vasileiou, K Evangelou, KG Vlasis… – Oxidative Medicine and …, 2020
The human thymus is a primary lymphoepithelial organ which supports the production of self-tolerant T cells with competent and regulatory functions. Paradoxically, despite the crucial role that it exerts in T cell-mediated immunity …
JM Brondello, YM Pers – Annals of Translational Medicine, 2019
… harboring shorten telomeres, by accumulating cycle-dependent kinases inhibitors driving senescence such as p16 Ink4a , p21 Cdkn1A and p57 KIP2 , and finally by producing deleterious catabolic and inflammatory mediators …
LW Harries – Role of RNA Modification in Disease, 2020
… For example, strategies are emerging now which allow selective delivery of senolytic cargoes to senescent cells only using galactosaccharide nanoparticles, which harness the observation that senescent cells harbor …
Ageing is characterized by the functional decline of tissues and organs and the increased risk of ageing-associated disorders. Several ‘rejuvenating’ interventions have been proposed to delay ageing and the onset of …
F Triana-Martínez, MI Loza, E Domínguez – Cells, 2020
… with the promise of future directions on innovative anticancer therapies. Keywords: cellular senescence; stemness; dormancy; quiescence; senolytic 1. Introduction Natural tumor evolution is a complex process, composed of …
NK September
… However, there is no single senolytic that has been shown to target all of our senescent cells … One example is Cleara Biotechnologies, whose founder, Dr. Peter De Keizer, has talked about senolytic “cocktails” and the problem …
B Cucarull, A Tutusaus, M Subías, M Stefanovic… – Cancers, 2020
… A-1331852 has been proposed as an agent in cancer therapy [42,43] and, more recently, as a senolytic compound [44]. Interestingly, through a dual mechanism acting on senescent cholangiocytes and activated fibroblasts …
M Demaria – Geriatric Oncology
… rejuvenation of aged-tissue stem cells (Childs et al. 2017; Soto- Gamez and Demaria 2017). Currently, a limited number of senolytic agents have been identified. 2-DG, a false substrate for the glycolytic metabolism, or bafilomycin A1, a
DE Citrin, PGS Prasanna, AJ Walker, ML Freeman…
… this setting. Although preventing premature senescence has shown promise in preventing RIF, there is increasing interest in agents that can clear prematurely senescent cells from tissues using “senolytic” drugs. Indeed, clearance …
S KOBASHIGAWA, YM SAKAGUCHI, S MASUNAGA… – Thermal Medicine, 2019
… In addition, a recent study demonstrated the efficacy of senolytic drugs in the selective elimination of senescent cells123); a combined treatment of dasatinib and quercetin (D+Q) was found to reduce the number of senescent …
MJ Yousefzadeh, J Zhao, C Bukata, EA Wade… – Aging Cell
Abstract Senescent cells accumulate with age in vertebrates and promote aging largely through their senescence‐associated secretory phenotype (SASP). Many types of stress induce senescence, includi…
N Martin, O Soriani, D Bernard – Trends in Molecular Medicine, 2020
The identification of senolytics, compounds that eliminate senescent cells, is presently a key priority given their therapeutic promise in cancer and aging-associated diseases. Two recent papers by Triana-Martínez et al. and Guerrero et al …
AJ Barinda, K Ikeda, DB Nugroho, DA Wardhana… – Nature Communications, 2020
Vascular senescence is thought to play a crucial role in an ageing-associated decline of organ functions; however, whether vascular senescence is causally implicated in age-related disease remains unclear. Here we show that …
BI Blog, ADN Booster
… Current research has shown that certain plant polyphenols (ie quercetin and fisetin) are strong senolytic agents (molecules that stop senescence), and have exciting potential for reducing and preventing senescence, which …
B Wang, Z Liu, VP Chen, L Wang, CL Inman, Y Zhou… – Aging Cell
… Our study potentially begins new avenues of research to discover whether pharmacological interventions, such as senolytic drugs (Tchkonia & Kirkland, 2018) or anti‐inflammatory drugs, can prevent or reverse dysfunction caused …
M Sharifi-Rad, R Pezzani, M Redaelli, M Zorzan… – Molecules, 2020
Page 1. Molecules 2020, 25, 467; doi:10.3390/molecules25030467 www.mdpi.com/ journal/molecules Review Preclinical Pharmacological Activities of Epigallocatechin- 3-gallate in Signaling Pathways: An Update on Cancer
K Kato, NJ Logsdon, YJ Shin, S Palumbo, A Knox… – American Journal of …, 2020
… One study demonstrated that the senolytic agent, quercetin, restored apoptosis susceptibility in IPF lung myofibroblasts, and inhibited fibrotic responses to lung injury in aged mice (62). Further, the first-in-human pilot study with a senolytic …
FJ Alibhai, F Lim, A Yeganeh, PV DiStefano… – Aging Cell
… Instead, we show that cellular senescence contributes to changes in particle cargo and function. Notably, senolytictreatment of old mice shifted plasma particle cargo and function toward that of a younger phenotype.
K Wolter, L Zender – Nature Reviews Gastroenterology & Hepatology, 2020
… cells. Therefore, the directed use of senolytic therapies in human cancers necessitates imaging modalities to non-invasively visualize TIS, thereby enabling image-guidedapplications of senolytic therapies. Full size image. Two …
MB Cavalcante, TD Saccon, ADC Nunes, JL Kirkland… – Aging, 2020
… Collagen deposition in the uterus is related to uterine aging. Senolytic therapies are an option for reducing health complications related to aging. We investigated effects of agingand the senolytic drug combination of dasatinib plus quercetin (D+Q) on uterine fibrosis …
C Martin-Ruiz, J Hoffmann, E Shmeleva, T von Zglinicki… – npj Aging and Mechanisms …, 2020
… p < 0.001; **p < 0.01; *p < 0.05 using 1-way ANOVA. Full size image. Senolytic therapy … The gating scheme is depicted in Fig. 2a. Animals, procedures and senolytictreatment. C57BL/6 mice were analyzed at either …
B Kang, Y Kim, TJ Park, HY Kang – Biochemical and Biophysical Research …, 2020
… allergic asthma. Br. J. Pharmacol., 173 (2016), pp. 1236-1247. Google Scholar. [8] JL Kirkland, T. Tchkonia, Y. Zhu, LJ Niedernhofer, PD RobbinsThe clinical potential of senolytic drugs. J. Am. Geriatr. Soc., 65 (2017), pp. 2297 …
PW O’Toole, PG Shiels – Journal of Internal Medicine, 2020
… Nrf2 regulates over 390 stress defence genes linked to longevity and health span. Moreover, alkyl catechols comprise a group of chemicals that include the senolytic agents fisetin and quercetin, which have proven efficacy in …
G Stolyarov II, A Grases
… When I am 85, the senolytic DNA machinery will be far from the only addition to my cells … Inflammatory conditions of aging will be a shadow of what they once were, because of senolytic therapies presently under development …
A Yabluchanskiy, S Tarantini, P Balasubramanian… – GeroScience
… Data are expressed as mean ± SEM. Results. WBI induces astrocyte senescence: protective effects of senolytictreatments … 3d, e). WBI impairs eicosanoid gliotransmitter-mediated NVC responses: protective effects of senolytictreatments …
CM Willis, AM Nicaise, ER Bongarzone, M Givogri… – Scientific Reports, 2020
The aging brain is associated with significant changes in physiology that alter the tissue microenvironment of the central nervous system (CNS). In the aged CNS, increased demyelination has been associated with astrocyte hypertrophy …
H Chen, H Chen, J Liang, X Gu, J Zhou, C Xie, X Lv… – Experimental & Molecular …, 2020
To study whether TGF-β1/IL-11/MEK/ERK (TIME) signaling mediates senescence-associated pulmonary fibrosis (SAPF) in Bmi-1-deficient (Bmi-1−/−) mice and determines the major downstream mediator of Bmi-1 …
MA Khan, X Wang, KTK Giuliani, P Nag, A Grivei… – International Journal of …, 2020
Proximal tubular epithelial cells (PTEC) are key players in the progression of kidney diseases. PTEC studies to date have primarily used mouse models and transformed human PTEC lines. However, the translatability of these …
BI Blog, HIA You
… Current research has shown that certain plant polyphenols (ie quercetin and fisetin) are strong senolytic agents (molecules that stop senescence), and have exciting potential for reducing and preventing senescence, which …
AH El-Far, NHE Darwish, SA Mousa – Integrative Cancer Therapies, 2020
Cellular senescence is a process of physiological growth arrest that can be induced by intrinsic or extrinsic stress signals. Some cancer therapies are associated with senescence of cancer cells wi…
M Ruiz-Ortega, S Rayego-Mateos, S Lamas, A Ortiz… – Nature Reviews Nephrology, 2020
Chronic kidney disease (CKD) is a devastating condition that is reaching epidemic levels owing to the increasing prevalence of diabetes mellitus, hypertension and obesity, as well as ageing of the population. Regardless …
L Yuan, PB Alexander, XF Wang – Science China Life Sciences, 2020
Cellular senescence (CS) is a state of stable cell cycle arrest characterized by the production and secretion of inflammatory molecules. Early studies desc.
H Li, MH Hastings, J Rhee, LE Trager, JD Roh… – Circulation Research, 2020
During aging, deterioration in cardiac structure and function leads to increased susceptibility to heart failure. The need for interventions to combat this age-related cardiac decline is becoming i…
X Li, J Wu, X Sun, Q Wu, Y Li, K Li, Q Zhang, Y Li… – Stem Cell Reports, 2020
JavaScript is disabled on your browser. Please enable JavaScript to use all the features on this page. Skip to main content Skip to article …
Y Li, J Lu, X Cao, H Zhao, L Gao, P Xia, G Pei – Oxidative Medicine and Cellular …, 2020
Oxidative stress-induced mitochondrial dysfunction and cell senescence are considered critical contributors to Alzheimer’s disease (AD), and oxidant/antioxidant imbalance has been a therapeutic target in AD. SIRT3 …